
Gene silencing is a general term describing epigenetic processes of gene regulation. The term gene silencing is generally used to describe the "switching off" of a gene by a mechanism other than genetic modification. That is, a gene which would be expressed (turned on) under normal circumstances is switched off by machinery in the cell.
Genes are regulated at either the transcriptional or post-transcriptional level.
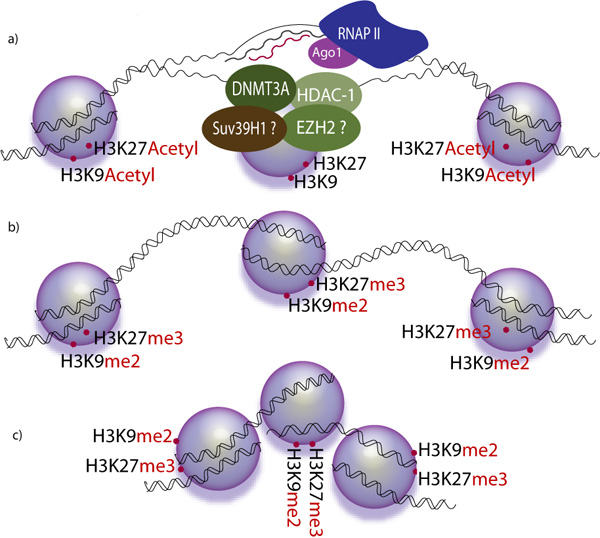
Transcriptional gene silencing is the result of histone modifications, creating an environment of heterochromatin around a gene that makes it inaccessible to transcriptional machinery (RNA polymerase, transcription factors, etc.).
Post-transcriptional gene silencing is the result of mRNA of a particular gene being destroyed. The destruction of the mRNA prevents translation to form an active gene product (in most cases, a protein). A common mechanism of post-transcriptional gene silencing is RNAi.
Both transcriptional and post-transcriptional gene silencing are used to regulate endogenous genes. Mechanisms of gene silencing also protect the organism's genome from transposons and viruses. Gene silencing thus may be part of an ancient immune system protecting from such infectious DNA elements.
RNA interference
The enzyme dicer trims double stranded RNA, to form small interfering RNA or microRNA. These processed RNAs are incorporated into the RNA-induced silencing complex (RISC), which targets messenger RNA to prevent translation.
RNA interference (RNAi) is a mechanism that inhibits gene expression by causing the degradation of specific RNA molecules or hindering the transcription of specific genes. The targets are often RNA from viruses and transposons (probably a form of innate immune response), although it also plays a role in regulating development and genome maintenance. Key to the RNAi processes are small interfering RNA strands (siRNA), which have complementary nucleotide sequences to a targeted RNA strand. The siRNA "guides" proteins within the RNAi pathway to the targeted messenger RNA (mRNA) and "cleaves" them, breaking them down into smaller portions that can no longer be translated into protein. A type of RNA transcribed from the genome itself, miRNA, works in the same way.
The selective and robust effect of RNAi on gene expression makes it a valuable research tool, both in cell culture and in living organisms because synthetic dsRNA introduced into cells can induce suppression of specific genes of interest. RNAi may also be used for large-scale screens that systematically shut down each gene in the cell, which can help identify the components necessary for a particular cellular process or an event such as cell division. Exploitation of the pathway is also a promising tool in biotechnology and medicine.
Cellular mechanism
The dicer protein from Giardia intestinalis, which catalyzes the cleavage of dsRNA to siRNAs. The RNase domains are colored green, the PAZ domain yellow, the platform domain red, and the connector helix blue.
RNAi is an RNA-dependent gene silencing process that is controlled by the RNA-induced silencing complex (RISC) and is initiated by short double-stranded RNA molecules in a cell's cytoplasm, where they interact with the catalytic RISC component argonaute.
dsRNA cleavage
Exogenous dsRNA initiates RNAi by activating the ribonuclease protein dicer, which binds and cleaves double-stranded RNAs (dsRNA)s to produce double-stranded fragments of 20–25 base pairs with a few unpaired overhang bases on each end.
Exogenous dsRNA is detected and bound by an effector protein, known as RDE-4 in C. elegans and R2D2 in Drosophila, that stimulates dicer activity. This protein only binds long dsRNAs, but the mechanism producing this length specificity is unknown. These RNA-binding proteins then facilitate transfer of cleaved siRNAs to the RISC complex.
This initiation pathway may be amplified by the cell through the synthesis of a population of 'secondary' siRNAs using the dicer-produced initiating or 'primary' siRNAs as templates. These siRNAs are structurally distinct from dicer-produced siRNAs and appear to be produced by an RNA-dependent RNA polymerase (RdRP).
RISC activation and catalysis
The active components of an RNA-induced silencing complex (RISC) are endonucleases called argonaute proteins, which cleave the target mRNA strand complementary to their bound siRNA. As the fragments produced by dicer are double-stranded, they could each in theory produce a functional siRNA. However, only one of the two strands, which is known as the guide strand, binds the argonaute protein and directs gene silencing. The other anti-guide strand or passenger strand is degraded during RISC activation. Although it was first believed that an ATP-dependent helicase separated these two strands, the process is actually ATP-independent and performed directly by the protein components of RISC.The strand selected as the guide tends to be that with a more stable 5' end, but strand selection is unaffected by the direction in which dicer cleaves the dsRNA before RISC incorporation. Instead, the R2D2 protein may serve as the differentiating factor by binding the less-stable 5' end of the passenger strand.
The structural basis for binding of RNA to the argonaute protein was examined by X-ray crystallography of the binding domain of an RNA-bound argonaute protein. Here, the phosphorylated 5' end of the RNA strand enters a conserved basic surface pocket and makes contacts through a divalent cation (an atom with two or more extra electrons) such as magnesium and by aromatic stacking (a process that allows more than one atom to share an electron by passing it back and forth) between the 5' nucleotide in the siRNA and a conserved tyrosine residue. This site is thought to form a nucleation site for the binding of the siRNA to its mRNA target.
Transcriptional silencing
Components of the RNA interference pathway are also used in many eukaryotes in the maintenance of the organisation and structure of their genomes. Modification of histones and associated induction of heterochromatin formation serves to downregulate genes pre-transcriptionally;this process is referred to as RNA-induced transcriptional silencing (RITS), and is carried out by a complex of proteins called the RITS complex. In fission yeast this complex contains argonaute, a chromodomain protein Chp1, and a protein called Tas3 of unknown function. As a consequence, the induction and spread of heterochromatic regions requires the argonaute and RdRP proteins. Indeed, deletion of these genes in the fission yeast S. pombe disrupts histone methylation and centromere formatiocausing slow or stalled anaphase during cell division. In some cases, similar processes associated with histone modification have been observed to transcriptionally upregulate genes.
The mechanism by which the RITS complex induces heterochromatin formation and organization is not well understood, and most studies have focused on the mating-type region in fission yeast, which may not be representative of activities in other genomic regions or organisms. In maintenance of existing heterochromatin regions, RITS forms a complex with siRNAs complementary to the local genes and stably binds local methylated histones, acting co-transcriptionally to degrade any nascent pre-mRNA transcripts that are initiated by RNA polymerase. The formation of such a heterochromatin region, though not its maintenance, is dicer-dependent, presumably because dicer is required to generate the initial complement of siRNAs that target subsequent transcripts.Heterochromatin maintenance has been suggested to function as a self-reinforcing feedback loop, as new siRNAs are formed from the occasional nascent transcripts by RdRP for incorporation into local RITS complexes. The relevance of observations from fission yeast mating-type regions and centromeres to mammals is not clear, as heterochromatin maintenance in mammalian cells may be independent of the components of the RNAi pathway.
Illustration of the major differences between plant and animal gene silencing. Natively expressed microRNA or exogenous small interfering RNA is processed by dicer and integrated into the RISC complex, which mediates gene silencing.
Biological functions
· Immunity
· Crosstalk with RNA editing
Technological applications
A normal adult Drosophila fly, a common model organism used in RNAi experiments.
An adult C. elegans worm, grown under RNAi suppression of a nuclear hormone receptor involved in desaturase regulation. These worms have abnormal fatty acid metabolism but are viable and fertile.
Gene knockdown
The RNA interference pathway is often exploited in experimental biology to study the function of genes in cell culture and in vivo in model organisms. Double-stranded RNA is synthesized with a sequence complementary to a gene of interest and introduced into a cell or organism, where it is recognized as exogenous genetic material and activates the RNAi pathway. Using this mechanism, researchers can cause a drastic decrease in the expression of a targeted gene. Studying the effects of this decrease can show the physiological role of the gene product. Since RNAi may not totally abolish expression of the gene, this technique is sometimes referred as a "knockdown", to distinguish it from "knockout" procedures in which expression of a gene is entirely eliminated.
Functional genomics
Most functional genomics applications of RNAi in animals have used C. elegans and Drosophila, as these are the common model organisms in which RNAi is most effective. C. elegans is particularly useful for RNAi research for two reasons: firstly, the effects of the gene silencing are generally heritable, and secondly because delivery of the dsRNA is extremely simple. Through a mechanism whose details are poorly understood, bacteria such as E. coli that carry the desired dsRNA can be fed to the worms and will transfer their RNA payload to the worm via the intestinal tract. This "delivery by feeding" is just as effective at inducing gene silencing as more costly and time-consuming delivery methods, such as soaking the worms in dsRNA solution and injecting dsRNA into the gonads. Although delivery is more difficult in most other organisms, efforts are also underway to undertake large-scale genomic screening applications in cell culture with mammalian cells.
Medicine
It may be possible to exploit RNA interference in therapy. Although it is difficult to introduce long dsRNA strands into mammalian cells due to the interferon response, the use of short interfering RNA mimics has been more successful. Among the first applications to reach clinical trials were in the treatment of macular degeneration and respiratory syncytial virus, RNAi has also been shown to be effective in the reversal of induced liver failure in mouse models.
Other proposed clinical uses center on antiviral therapies, including the inhibition of viral gene expression in cancerous cells, knockdown of host receptors and coreceptors for HIV, the silencing of and hepatitis B genes, silencing of influenza gene expression, and inhibition of measles viral replication.Potential treatments for neurodegenerative diseases have also been proposed, with particular attention being paid to the polyglutamine diseases such as Huntington's disease.RNA interference is also often seen as a promising way to treat cancer by silencing genes differentially upregulated in tumor cells or genes involved in cell division A key area of research in the use of RNAi for clinical applications is the development of a safe delivery method, which to date has involved mainly viral vector systems similar to those suggested for gene therapy.
Biotechnology
RNA interference has been used for applications in biotechnology, particularly in the engineering of food plants that produce lower levels of natural plant toxins. Such techniques take advantage of the stable and heritable RNAi phenotype in plant stocks. For example, cotton seeds are rich in dietary protein but naturally contain the toxic terpenoid product gossypol, making them unsuitable for human consumption. RNAi has been used to produce cotton stocks whose seeds contain reduced levels of delta-cadinene synthase, a key enzyme in gossypol production, without affecting the enzyme's production in other parts of the plant, where gossypol is important in preventing damage from plant pests. Similar efforts have been directed toward the reduction of the cyanogenic natural product linamarin in cassava plants.
Conclusion
Gene silencing is a general term describing epigenetic processes of gene regulation. The term gene silencing is generally used to describe the "switching off" of a gene by a mechanism other than genetic modification. That is, a gene which would be expressed (turned on) under normal circumstances is switched off by machinery in the cell. Transcriptional gene silencing is the result of histone modifications, creating an environment of heterochromatin around a gene that makes it inaccessible to transcriptional machinery (RNA polymerase, transcription factors, etc.).